Эволюция как сопротивление энтропии
Виктор Прохорович Щербаков
доктор биологических наук,зав. лабораторией молекулярной генетики Института проблем химической физики РАН
I. Механизмы видового гомеостаза
Организм и среда
Что отбирает отбор?
Гомеостаз
Об адаптивном мутагенезе и эволюции способности к эволюции
Совершенство
Генетический «замысел» зиготы
II. Консервативная роль полового размножения
Когезия
Генетическая рекомбинация
Необратимость эволюции. Храповик Мюллера
Репродуктивная изоляция
Список литературы
Эволюция направлена против себя самой
Эволюция – это способ живого противостоять энтропии, нарастающему хаосу и беспорядку. Она творит разные новшества, но естественный отбор сохраняет только те из них, что придают организмам устойчивость к дальнейшим изменениям, те, что позволяют им воспроизводить свои копии в длинном ряду поколений, практически не меняясь. Как это ни странно, но получается, что эволюция работает против себя самой.
Мы привыкли к тому, что эволюция есть создание чего-то нового, более сложного и совершенного. Но на самом деле эволюция – это создание не просто нового, а нового, которое сопротивляется дальнейшим изменениям. Иными словами, эволюция направлена на прекращение эволюции! К такому парадоксальному выводу пришел Виктор Прохорович Щербаков из Института проблем химической физики РАН (Черноголовка, Московская область). В своей статье, опубликованной в двух номерах «Журнала общей биологии», он подробно обсуждает и детализирует этот тезис, опираясь прежде всего на литературу в области молекулярной биологии.
Суть жизни Щербаков видит в противостоянии энтропии, растущему хаосу. Удивительно то, что, сопротивляясь энтропии, эволюция на самом деле этой самой энтропией движима. Так, никуда не деться организмам от мутаций – сбоев в механизме передачи наследственной информации от родителей к потомкам. Мутации приводят в конце концов к гибели организмов и вымиранию видов. Но удивительно то, что в ходе этого разрушительного по сути своей процесса (частного проявления энтропии) случайно создаются новшества, которые опять же случайно могут оказаться устойчивыми к дальнейшей деградации. Вот они-то и сохраняются отбором. Так возник когда-то генетический код (недаром он универсальный для всех организмов!) и механизм воссоздания организмами своих копий из материала окружающей среды, так появился диплоидный набор хромосом и половое размножение (смысл которого автор видит в стабилизации вида, в создании препятствий на пути его дальнейшей эволюции), так возникала забота о потомстве и разные другие сложнейшие формы поведения животных (а в конце концов и наша культура). Короче говоря, так сформировалось всё то, что позволяет организмам воспроизводить себя в потомках, не исчезая с лица Земли.
Палеонтологи часто жалуются на то, что не могут найти форм переходных, промежуточных между разными группами организмов. Но с развиваемой точки зрения это неудивительно. Переходные формы не сохранились, поскольку не были устойчивыми, поскольку вынуждены были претерпевать быструю эволюцию, и только ценой существенного изменения своего строения (и/или физиологии) могли сохранять соответствие среде (которая сама менялась), могли выживать и оставлять жизнеспособных потомков. Но на каком-то этапе в этом ряду форм, быстро меняющихся из поколения в поколение, появились организмы, более совершенные, лучше приспособленные к среде, к тому, чтобы жить в этой среде не меняясь. Такие более совершенные формы уже могли сохранить свою собственную идентичность в длинном ряду поколений. А поскольку они долго пребывали на Земле, то и остатки их (отпечатки, скелеты и т. п.) обнаруживались учеными достаточно часто. Впрочем, эта точка зрения не нова. Она была очень четко сформулирована в 1972 году Нильсом Элдриджем (Niles Eldredge, р. 1943) и Стивеном Гоулдом (Stephen Jay Gould, 1941–2002) в виде гипотезы прерывистого равновесия (punctuated equilibrium).
Алексей Гиляров
I. Механизмы видового гомеостаза
Живой организм – это временный материальный носитель организации
со свойством выживания.
Е. Г. Мерсер (Merser, 1981)
Второй закон – это больше, чем закон термодинамики,
это естественный закон истории.
Д. Р. Брукс и Е. О. Вили (Brooks, Wiley, 1986)
Подобно тому, как в зерне невидимо содержится всё, что должно со временем
развиваться в дерево, так следует нам представлять себе,
что и мир в момент, когда Бог одновременно сотворил все вещи,
содержал в себе все вещи, которые земля произвела,
как возможности и как причины, прежде чем они развились
во времени такими, какими их знаем мы.
Блаженный Августин
Слово «эволюция» ассоциируется с образованием нового. Проблема эволюционной стабильности редко привлекает к себе внимание эволюционистов. Чаще всего, в качестве курьезов, упоминаются эволюционные долгожители – живые ископаемые (гаттерия, крокодилы, устрицы, мечехвост, опоссум, гинкго), хотя в последние два десятка лет, с появлением теории прерывистого равновесия, механизмы стазиса начинают обсуждать в позитивном ключе. В данном очерке сделана попытка обосновать точку зрения, согласно которой онтологическим содержанием биологической эволюции является не просто создание новых форм, а форм, устойчивых к дальнейшей эволюции, что в долговременном плане, в геологических масштабах времени сохраняются только устойчивые к эволюции формы. Рассматривая эволюцию как энтропийный процесс, мы можем говорить о создании форм, противостоящих росту энтропии. Ныне существующие и известные палеонтологам виды организмов представляют собой выборку долгожителей из общей массы когда-либо возникавших форм. Это представление может в какой-то мере объяснить отсутствие переходных форм в современной биосфере и неполноту палеонтологической летописи тем, что быстро эволюционирующие виды наряду с быстро вымирающими видами не оставляют заметных следов в истории биосферы.
Организм и среда

Крокодил – это самое настоящее живое ископаемое. Плавающие рядом рыбы из семейства цихловых – сравнительно молодые создания. А что уж говорить о человеке! Этот снимок румынского фотографа Джордже Хренюка (George Hreniuc) так и называется: «Эволюция». (Фото с сайта codedlanguage.org)
Теория естественного отбора ввела эволюционную биологию в русло естественнонаучного мышления, освободив ее от креационистских взглядов, витализма и веры в наследование приобретенных признаков («бегство от чуда»). Заблуждение ламаркизма коренится в неверном взгляде на взаимоотношение организма и среды, а именно в представлении о формировании организма средой. Это представление иногда присутствует даже в работах авторов, которые сознательно не разделяют ламаркистских верований, в виде таких формулировок, как «фенотип есть результат взаимодействия генотипа со средой», «свойства организма равно зависят от генотипа и среды, как площадь прямоугольника зависит от ширины и длины» и т. п. Особенно выпукло эта путаница видна в известном понятии «наследование» признака. В выражении типа «данный признак имеет наследование 80%» не подразумевается же, что признак на 20% может быть без генов? К сожалению, даже и прочно укоренившееся выражение «приобретенные признаки» некорректно и ведет к недоразумениям. Во-первых, все признаки, конечно же, являются приобретенными, и все они не наследуются, потому что наследуется, строго говоря, оплодотворенная яйцеклетка с ее хромосомами и органеллами, белками и мембраной. Всё остальное приобретается в ходе онтогенеза. Онтогенез идет, разумеется, в окружающей среде, но участники пары «организм—среда» не изоморфны и не могут сопоставляться как равные партнеры.
Всякий знакомый с молекулярной и клеточной биологией знает, как сложна, упорядоченна и высокоорганизованна даже простейшая клетка. Ни одно из наших технологических достижений, включая суперкомпьютеры и космические корабли, не выдержит сравнения с организацией живой клетки. Клетка – подлинное термодинамическое чудо, крайне маловероятная система. А что такое среда? Например, для цианобактерий и всех зеленых растений минимальная среда включает воду, углекислый газ, несколько минеральных солей и свет. В такой среде клетка живет, размножается, строит себя, синтезируя при этом тысячи разных белковых молекул и других органических веществ, включая нуклеиновые кислоты и массу низкомолекулярной органики. Сотворение мира из почти ничего! Среда хаотична, организм упорядочен и высокоорганизован; организм созидает, среда деструктивна; организм в своем жизненном цикле осуществляет генетический «замысел», среда не имеет никакого замысла об организме; организм «знает» среду, избирательно берет из нее то, что ему нужно, и защищается от того, что ему вредно или опасно, среда ничего не знает об организме; организм живет, а среда мертва. В структурном, термодинамическом и информационном отношении организм неизмеримо выше среды. Организм есть сущее, а среда – условия, в которых организм существует. К компонентам среды относится, разумеется, и энергия.
Выражение «среда модифицирует организм» эвристически вредно. Это организмы обладают способностью к приспособительному изменению своего фенотипа в разных условиях среды без изменения генотипа. Эта способность, известная уже у бактерий, – одно из важнейших достижений эволюции. Как и все прочие свойства организма, она определяется генами.
Дарвиновский отбор случайных и редких наследственных изменений выводит организмы из-под размывающего действия среды, поскольку сами изменения не определяются средой. Недостаточность синтетической теории в качестве общей теории биологии осознается многими. Я, впрочем, не разделяю часто высказываемый упрек дарвинизму в неспособности предсказывать ход эволюции. Как всякий творческий процесс, эволюция несет в себе момент неопределенности и неповторимости. Понимать эволюцию можно только ретроспективно. Если следовать Карлу Попперу (Popper, 1965), наука должна ограничиться очень простыми явлениями, воспроизводимыми и обратимыми. Все творческие процессы должны быть, следовательно, исключены как объекты исследования. В этом случае, конечно, законы природы сводятся к законам физики. Не биологические теории плохи – не умеют предсказывать, – а «плоха» природа – непредсказуема.
Механизм адаптивной эволюции в его предельном (не реальном) случае, когда селективная ценность новых аллелей приближается к единице, становится неотличимым от ламаркистского по своим последствиям. Быстро адаптируясь к меняющимся условиям, организмы и виды утрачивают самоидентичность, становятся функцией среды. Реальные виды, однако, устойчивы во времени и, следовательно, оказывают сопротивление средовым воздействиям, не только прямым (ламаркизм) но и в качестве отбирающего агента (дарвинизм). Адаптационная эволюция, не будь она жесточайше ограничена необходимостью сохранить внутреннее совершенство организма, вела бы к тому же, что и ламаркистская эволюция, – к деградации и хаосу, к броуновскому движению, к эфемеризации видов, низведению организмов до термодинамического уровня среды. В этом отношении виртуальный термин «приспособленность», характеризующий успешность размножения особи, эвристически неудачен, так как легко понимается как «приспособленность к среде». О примате организма над средой и о теоретической важности проблемы закрытости биологических систем см. статью Брукса (Brooks, 2000).
Что отбирает отбор?
Существует всеобщее убеждение в самоценности биологической эволюции, понимаемой исключительно как создание новшеств. Уже по меньшей мере полтора столетия слова «прогресс», «эволюция» воспринимаются однозначно позитивно. Только этой общей направленностью умов можно объяснить малое внимание к тому, что онтологическое содержание эволюции – это не просто сотворение новых форм, но сотворение форм, устойчивых к дальнейшим изменениям, что объективно дело выглядит таким образом, как будто эволюция борется против дальнейшей эволюции.
Неодарвинистская парадигма эволюции – отбор случайных наследственных вариантов по признаку приспособленности или селективное размножение генотипов в соответствии с приспособленностью всегда подвергалось критике как за тавтологию (выживание выживающих), так и за то, что она оставляет в стороне движущие силы эволюции, ее отношение к общим законам природы, в частности термодинамическим, и не дает объяснения очевидной временной направленности эволюции и ее необратимости. Если механизм эволюции состоит в отборе всё более приспособленных форм, то должна наблюдаться «стрела приспособленности». Однако мы не можем даже утверждать, что человек более приспособлен, чем бактерия.
Попытки связать приспособленность с термодинамическим принципом минимального производства энтропии (Prigogine et al., 1972; Hamilton, 1977) выглядят обнадеживающими, хотя чисто физическую эффективность использования энергии едва ли удастся применить в качестве той величины, которая непрерывно возрастает в ходе эволюции, подобно тому как во всех физико-химических процессах в природе возрастает энтропия. Теплокровные животные и в особенности человек с его энергетическим расточительством явно выпадают из этой закономерности.
Сондерс и Хо (Saunders, Ho, 1976, 1981) по аналогии с принципом минимального производства энтропии постулируют принцип минимального возрастания сложности в эволюции. «Сложность», «упорядоченность», «организованность» часто используются в литературе по эволюции и не всегда в одинаковом значении, чаще всего в соответствии с интуитивным пониманием авторами этих слов. В теории информации, рассматривающей физическую систему как последовательность цифр, необходимую для ее описания (Kolmogorov, 1968; Chaitin, 1974, 1975), сложность определяется как информационное содержание кратчайшего алгоритма, достаточного для однозначного воспроизведения этой последовательности. Случайная или беспорядочная последовательность характеризуется в этой теории как максимально сложная, несжимаемая (Chaitin, 1975), то есть она может быть определена только цифра за цифрой, поскольку в ней полностью отсутствует упорядоченность, которая позволяла бы описать ее в виде более краткого алгоритма. Упорядоченная последовательность, следовательно, может быть сжата до алгоритма, информационное содержание которого меньше такового самой последовательности.
Эти концепции в общем близки к интуитивным понятиям. В определении организованности я буду придерживаться концепции Денбая (Denbigh, 1975), в которой постулируется, что организованная система – это сложная система, обладающая определенной функцией благодаря наличию специфических связей между элементами системы. Организованные системы следует отличать от упорядоченных. И те, и другие не являются случайными, но если упорядоченные системы могут быть генерированы с помощью простых алгоритмов и, следовательно, лишены сложности, организованные системы должны быть собраны элемент за элементом в соответствии с внешней программой или замыслом. Организация, следовательно, есть сложность, наделенная функцией. Она неслучайна в результате интеллектуального конструирования или естественного отбора, а не из-за априорной необходимости кристаллографического порядка (Wicken, 1979).
Движение к большей сложности – очевидная общая тенденция эволюции биосферы. Однако сама сложность не означает ни более высокой приспособленности, ни какого-либо иного совершенства. Напротив, если иметь в виду задачу выживания, сохранения, сложное более чувствительно к деградации, чем простое. Используя «геометрическую модель» Фишера (Fisher, 1930), Ор (Orr, 1998; 1999; 2000) и позднее Вэлч и Ваксман (Welch and Waxman, 2003) показали, что увеличение числа фенотипических признаков коррелирует со снижением скорости адаптации. Ор назвал это «ценой сложности». Стабильность существующих сложных систем критически зависит от наличия специфических, неслучайных связей между ее частями (Saunders, Ho, 1976).
Почему естественный отбор отбирает сложное? Может быть, это и не так. Наряду с появлением более сложного эволюция сохраняет и даже творит заново и более простые формы. Сложные отнюдь не вытесняют простых. Есть человек, но существуют (и процветают!) бактерии. Вся естественная история наличествует в сегодняшней природе (кроме гипотетических доклеточных форм жизни). Так что «стрела сложности» связана, по крайней мере отчасти, с тем, что эволюция по необходимости начинала с простого. Более древняя эволюция простых форм в большой мере уже себя исчерпала (достигла совершенства), и жизнеспособное новое возникает преимущественно на путях усложнения.
Принцип селекции стабильных структур является общим для добиотической и биологической эволюции. Объекты Вселенной имеют очень разные времена жизни, от ничтожных долей секунды до миллиардов лет. В ходе эволюции идет замена эфемерных форм на более стабильные: сохраняется лишь то, что долго сохраняется. В живых системах сопротивление гибели достигается тем, что в ходе биологической эволюции сохраняется лишь то, что не изменяется.
Одна из формулировок второго закона термодинамики звучит так: все системы самопроизвольно меняются таким образом, что уменьшается их способность к изменению, то есть они стремятся к состоянию равновесия. Живые организмы – термодинамически неравновесные системы, устроенные таким образом, что, пока они живы, они стремятся не к равновесию, а к некоторому неравновесному, но устойчивому состоянию, называемому стационарным. Механизмы, обеспечивающие стационарное состояние, называются гомеостазом. Энергия, будучи фактором хаотическим, сама по себе недостаточна для удержания системы в состоянии, далеком от равновесия, – необходимо, чтобы эта система была определенным образом устроена, она должна быть диссипативной структурой.
Гомеостаз

Это тоже своего рода равновесие. Рисунок Маурица Корнелиса Эшера (1898–1972). (С сайта www.enseignement.polytechnique.fr)
Развитие и последующая жизнь индивида идут под управлением генетических программ, обеспечивающих целенаправленное использование энергии и веществ окружающей среды для построения своего тела, поддержания его в стационарном состоянии и оставления потомства. Я буду использовать выражение «под управлением генов» ради удобства, хотя мы всегда должны помнить, что сами по себе гены – текст, приобретающий смысл только в контексте организма. Несмотря на видимую гибкость генетических инструкций и пластичность в отношении средовых эффектов, конечная цель онтогенеза достигается с поразительной точностью. Это означает, что онтогенез в своих существенных чертах – детерминированный процесс. Это не механическая детерминированность, а детерминированность стохастическими средствами. Биологический детерминизм условен в том смысле, что результат в определенной мере зависит от внешних условий. Тем не менее это детерминизм, так как в определенных условиях результат будет определенным. Вторая сторона условности биологического детерминизма в том, что генетический «замысел» об организме может быть раскрыт только эмпирически и не может быть вычислен исходя из начальных условий. Предопределено, но непредсказуемо. Случаи пороков развития и вызванных внешними условиями уродств, как и абортивное развитие, – следствие либо несовершенства данного конкретного генома, либо наличия таких средовых факторов, которые находятся за пределами адекватной реакции генетической программы.
Итак, организмы способны противостоять стремлению к хаосу, противополагая ему умение использовать энергию и другие компоненты внешней среды для поддержания своих структур и функций, то есть в конечном счете для сохранения себя во времени. Организмы, однако, представляют собой сложные системы, далекие от максимума энтропии. Они энтропийно напряжены. Механизмы гомеостаза не являются совершенными, поэтому ни один индивидуальный организм не может избежать термодинамического равновесия, то есть смерти. Организмы слишком сложны, слишком маловероятны, чтобы долго выдерживать давление энтропии. Простые атомы и молекулы существуют почти столько же, сколько существует Вселенная, а жизнь индивидуального организма конечна.
Отчего гибнут организмы? Концепция естественного отбора и борьбы за существование настолько овладела нашим сознанием, что мы не придаем важности тому, что при самых благоприятных условиях жизни, в отсутствие всякой конкуренции, при изобилии источников энергии и вещества организмы всё равно неизбежно погибают. Они погибают от энтропии. Живые системы обходят термодинамический запрет с помощью размножения, то есть копирования самих себя, снятия реплик. Иногда одноклеточные организмы в отличие от многоклеточных рассматривают как бессмертные. Это недоразумение. Они тоже спасаются размножением.
Каждая индивидуальная реплика существует сравнительно недолго, но виды потенциально бессмертны и фактически некоторые существуют неизменными десятки и сотни миллионов лет, а докембрийские организмы, жившие три миллиарда лет назад, видимо, мало отличались от современных цианобактерий (Fox, Dose, 1972). На обратную сторону процесса эволюции – сохранение уже существующей, сформировавшейся в ходе предшествующей эволюции организации биологических систем, стасигенез или стазис, — обращали внимание многие эволюционисты (Huxley, 1957; Шмальгаузен, 1968; Dobrzhansky, 1970; Gould, Eldredge, 1977; Расницын, 1986; Северцов, 1986, 1987, 1990). Однако в увлечении идеей прогрессивного хода эволюции мы не склонны придавать значение тому, как мало меняются организмы, как они на самом деле устойчивы к эволюции. Весь современный органический мир состоит из эволюционных долгожителей. Человек среди них самый молодой, но и нам уже может быть миллион лет.
Копирование, репликация – не совсем точные слова. Реплицируются только гены, а организмы воспроизводятся de novo в соответствии с генетическим «замыслом» (см. ниже). Вместо безнадежного дела сохранения сложных материальных структур организма сохраняется информация о нём. Это гораздо более простая задача – организм несравненно сложнее (и значит, уязвимее для энтропии), чем его ДНК. Хранение информации, а не тел – важнейший атрибут живого и неодолимый довод против возможности сведения биологии к физико-химии. Всякое кодирование связано с использованием символов, но символ связан с символизируемым не физико-химически, а семантически. Здесь – параллель между аналоговыми и цифровыми системами связи. Замена одной пары оснований, скажем, пары АТ на пару ТА в молекуле ДНК, содержащей миллиарды таких пар, ничего собой не представляет ни с точки зрения физико-химии, ни с точки зрения теории информации. А между тем указанная замена может оказаться смертельной, приведет к гибели весь организм, сложную, высокоорганизованную систему.
Биологические виды намного более стабильны, чем индивидуальные организмы. Более того, мы не знаем принципиальных препятствий для их вечного существования при условии сохранения адекватной окружающей среды. Виды также представляют собой системы, далекие от равновесия, и потому должны обладать механизмами гомеостаза. Именно в этом ключе, с позиции совершенствования гомеостаза вида, нужно рассматривать эволюцию (Slobodkin, Rapoport, 1974).
Размножение организмов позволяет им избежать неизбежного, казалось бы, распада и сохраниться во времени, несмотря на свою сложность, на температуру, превышающую 300° и агрессивную окружающую среду, причем организмы не находятся в состоянии консервации, а активно живут в этих условиях. Главный секрет этого термодинамического чуда в самом размножении, то есть в получении тысяч, миллионов, миллиардов копий генетической информации. Любой онтогенез, любой жизненный цикл имеет своим фокусом получение этих копий. На этой задаче сосредоточены в конце концов все системы организма и все его затраты. Копирование, однако, сопряжено с возможностью ошибок, а копирование с ошибками не решает задачу создания совершенного гомеостаза вида.
Точность копирования генетического материала. Это первый и главный антиэволюционный барьер, созданный эволюцией. Теоретически ясно, что точность репликации должна быть такой, чтобы большинство потомков получали не содержащую ошибок генетическую информацию. Наиболее надежные измерения скорости мутирования выполнены на микроорганизмах, как прокариотических, так и эукариотических. Точность репликации ДНК оказалась поразительно высокой. Например, скорость спонтанных мутаций в растущих клетках кишечной палочки составляет всего лишь около 0,003 ошибки на геном на одну репликацию ДНК (Drake, 1991; Drake et al., 1998). Это около 6 × 10–10 мутаций на пару оснований ДНК. Другие микроорганизмы, в том числе эукариотические, имеют не менее высокую точность репликации (таблица).
Точность копирования ДНК у микроорганизмов
(Drake, 1991; Drake et al., 1998)
| Организм | Размер генома (нуклеотиды) |
Частота мутаций | |
| на пару нуклеотидов | на геном | ||
| Бактериофаг М13 | 6,4 × 103 | 7,2 × 10–7 | 0,0046 |
| Бактериофаг λ | 4,9 × 104 | 7,7 × 10–8 | 0,0038 |
| Бактериофаги Т2 и Т4 | 1,7 × 105 | 2,4 × 10–8 | 0,0040 |
| Escherichia coli | 4,6 × 106 | 5,4 × 10–10 | 0,0025 |
| Saccharomyces cerevisiae | 1,2 × 107 | 2,2 × 10–10 | 0,0027 |
| Neurospora crassa | 4,2 × 107 | 7,2 × 10–11 | 0,0030 |
| В среднем | 0,0034 | ||
Интересно, что самые разные организмы – бактериофаги,
бактерии, грибы – дают близкие величины числа мутаций на геном на
репликацию, хотя размер генома (и соответственно скорость мутирования на
пару оснований) варьируют в пределах четырех порядков величины. Это
показывает, что эволюционно значимым параметром является скорость
мутирования в расчете на геном на поколение. Для одноклеточных
организмов достигнутой высокой точности копирования и эффективности
репарации ДНК достаточно для надежного превышения скорости размножения
над скоростью мутирования (так, чтобы подавляющее большинство потомков
не имело ни одной новой мутации) и обеспечения, таким образом,
потенциального бессмертия вида.
У многоклеточных организмов (Drake et al., 1998; Crow, 1997) точность синтеза ДНК приблизительно такая же, как у одноклеточных эукариот (до 5 × 10–11 на пару оснований), однако число мутаций на геном на половое поколение оказывается очень высоким. Например, у человека число новых мутаций на зиготу составляет более 60. Трудно себе представить, как такое размывающее действие мутагенеза совместимо с жизнью. Проблема связана с огромными размерами генома у высших эукариот и большим числом клеточных делений в зародышевом пути (Crow, 1997).
Одно из объяснений переносимости столь высокого уровня мутагенеза у высших организмов состоит в предположении, что огромная доля их генома – это не функциональные гены, а внутригенные вставки (интроны) и межгенные последовательности, мутации в которых нейтральны или почти нейтральны. Пересчет числа мутаций на половое поколение на «эффективный геном» дает величины от 0,036 у мелкой нематоды Caenorhabditis elegans до 1,6 у человека (Drake et al., 1998) – всё еще угрожающе высокий груз мутаций. Селективная нейтральность большинства мутаций (следовательно, и их практическая безвредность) была даже положена в основу теории молекулярной эволюции (Кимура, 1985). Я, впрочем, думаю, что нейтральные изменения вообще не представляют собой эволюцию. Практическая нейтральность мутаций означает только то, что так называемая молекулярная эволюция уже в основном завершена, и мы видим только вариации, не имеющие эволюционного значения (вырожденность связи структура—функция).
Цена точности. Почему скорость мутирования не падает до нуля? Дело, по-видимому, в том, что в конце концов стремление к увеличению точности копирования генов оказывается уравновешенным всё возрастающей ценой этой точности (Kimura, 1967; Kondrashov, 1995). Это следует и из общих термодинамических соображений: очевидно, что для достижения абсолютной точности понадобились бы бесконечные энергетические затраты. Уже достигнутая точность копирования – одна ошибка на десятки миллиардов знаков – очень высока. Сложные и разнообразные механизмы сохранения и копирования генетической информации находятся в ряду главных достижений биологической эволюции, но их функционирование является и главной статьей энергетических затрат клетки (Cox, 1994). Представляется вероятным, что достигнутая у высших организмов точность синтеза ДНК и эффективность репарации ее повреждений близки к предельным. И это означает, что организмы с большим размером генома должны обладать дополнительными механизмами, позволяющими виду сохраняться в условиях сильного мутационного давления.
Очищающий отбор. Термин «усеченный (или отсекающий) отбор» (truncated selection) обычно используют для обозначения однонаправленной селекции, когда для размножения отбирают особей с количественным признаком выше или ниже определенной пороговой величины. Позднее этим термином стали обозначать также отбор, при котором отбраковываются особи с мутационным грузом выше определенной пороговой величины. Это отбор на сохранение дикого генотипа, противоположный однонаправленному. Во избежание путаницы я буду называть его очищающим отбором. Мутагенез – случайный процесс, и число новых мутаций, приобретаемых потомками, варьирует в соответствии с пуассоновым распределением. Иначе говоря, среди потомков всегда присутствуют особи с числом мутаций меньше и больше среднего. Поскольку мутации чаще всего вредны, то есть снижают жизненность, а вредный эффект мутаций может быть аддитивным или даже синергическим (положительный эпистаз), особи с повышенным числом мутаций будут иметь низкую жизненность, тогда как особи с малым числом мутаций или без таковых получают преимущества. Формула «побеждает наименее отягощенный мутациями» более точна, во всяком случае, более конкретна, чем «побеждает сильнейший». Так идет очищение популяции от мутационного груза, но популяция при этом платит определенную цену, производя нежизнеспособное потомство. Цена тем выше, чем выше скорость мутагенеза. Здесь тоже ожидается предельная мутагенная нагрузка, выше которой цена очищающего отбора становится несовместимой с выживанием вида. Эффективность очищающего отбора существенно возрастает, а цена его падает при наличии полового размножения и генетической рекомбинации (Crow, Kimura, 1979; Kondrashov, 1982, 1988).
Стабилизирующий отбор. В популяциях, хорошо адаптированных к условиям обитания, действует стабилизирующий, или нормализующий, отбор (Шмальгаузен, 1968), при котором уклоняющиеся варианты оказываются менее приспособленными, чем «средний» фенотип.
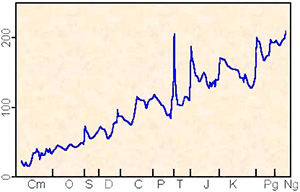
Яркая иллюстрация идеи В.П.Щербакова о неизбежном замедлении эволюции – рост средней продолжительности существования родов морских животных в течение последних 540 млн лет, подтверждаемый данными палеонтологии. По горизонтальной оси: геологическое время; по вертикальной оси: средняя продолжительность существования родов морских животных.
Рис. из статьи А.В.Марков, 2000
Пределы адаптационной эволюции. Есть важная сторона эволюции, автоматически снижающая возможность дальнейшей эволюции. Общей предпосылкой неизбежного замедления эволюции является то, что организм представляет собой целостную, интегрированную, когерентную систему, в которой любое изменение так или иначе нарушает общую отлаженность и согласованность систем организма и либо должно быть отброшено отбором, либо компенсировано другими изменениями (отрицательные обратные эволюционные связи) (Kauffman, 1973; 1983; Zuckerkandl, 1976; Brooks, Wiley, 1986; Robertson, 1991; Seaborg, 1999). Чем более совершенны онтогенетические программы организма, тем менее вероятно, что случайная мутация может их улучшить, и в пределе мы должны ожидать полной невозможности дальнейших позитивных изменений подобно тому, как в стихотворении, написанном гением, нельзя изменить ни одного слова без того, чтобы его не ухудшить. Здесь потребности адаптации к меняющимся условиям среды сталкиваются с необходимостью удержать достигнутое внутреннее совершенство. Чем важнее система для внутреннего совершенства, тем менее способна она к изменению. Примером такой стабильности является универсальность генетического кода во всём живом мире. Возможно, впрочем, что дело не во внутреннем совершенстве генетического кода (интуитивно кажется, что могли бы быть и другие, не менее совершенные варианты кода). Может быть, само появление полноценного генетического кода поставило его владельца вне конкуренции и сделало родоначальником всей биосферы (эффект основателя). Весьма консервативны все фундаментальные генетические процессы: синтез, репарация и рекомбинация ДНК, транскрипция, синтез белка. Мутации в них должны быть крайне вредными или летальными.
Устойчивому существованию вида грозят не только вредные мутации, но и «полезные», потенциально способные превратить один вид в другой, что для данного вида означает исчезновение. Быстро эволюционирующий вид становится жертвой «приспособленчества». В пределе должен быть достигнут такой уровень совершенства, когда полезные мутации становятся невозможными.
Диплоидия. Для высших организмов, растений и животных, характерен жизненный цикл со сменой гаплоидной и диплоидной фаз, причем протяженная гаплоидная фаза характерна для организмов с простым развитием, тогда как для организмов со сложным онтогенезом (многоклеточные животные и семенные растения) характерна протяженная диплоидная фаза. У высших животных гаплоидными являются только гаметы. Существуют различные (не исключающие друг друга) точки зрения на механизмы происхождения и поддержания диплоидного состояния (Crow, Kimura, 1965; Charlesworth, 1991; Kondrashov, Crow, 1991; Perrot et al., 1991; Goldstein, 1992; Orr, 1995). Для обсуждаемой здесь проблемы существенным является общее соображение, что диплоидия может представлять собой путь повышения долговечности вида. У гаплоидов генетическая информация хранится в двух копиях, на двух комплементарных нитях ДНК, у диплоидов – в четырех копиях, что, разумеется, повышает надежность ее сохранения. Мутации в двух наборах хромосом редко затрагивают одни и те же сайты, так что всегда сохраняется принципиальная возможность восстановления исходного текста. Практическая возможность такого восстановления существует у видов с половым размножением, и значение диплоидности следует рассматривать в контексте полового размножения и рекомбинации. Отметим, что гаплоидный геном характерен как раз для организмов, размножающихся бесполо, а облигатное половое размножение у высших животных неразрывно связано с их диплоидностью. Корреляция между сложностью развития и большим геномом, с одной стороны, и преобладанием диплоидной фазы – с другой понятна. У одноклеточного организма нет проблемы избавления от мутационного груза, поскольку большая часть потомков не несет ни одной мутации. Но эта проблема существует у организмов с большим геномом и длинным зародышевым путем.
Кроме увеличения надежности сохранения генов в поколениях, диплоидность делает существенный вклад в сохранение индивидов в ходе онтогенеза (Orr, 1995), который важен и для половых, и для бесполых видов. Большая часть новых мутаций в той или иной мере рецессивна, и диплоидность резко повышает толерантность к мутациям, как к унаследованным от родителей, так и к соматическим (Crow, Kimura, 1965; Kondrashov, Crow, 1991; Drake, 1991; Orr, 1995). Выгода не ограничивается маскированием рецессивных мутаций. Диплоидность обеспечивает возможность рекомбинационной репарации повреждений в ДНК, в том числе таких, которые не репарируются более простыми системами репарации.
Существенным фактором повышения жизнеспособности вида в результате диплоидности является преимущество гетерозигот (Lerner, 1954; Goldstein, 1992). Полиморфные популяции свободно скрещивающихся организмов обычно более устойчивы к средовым влияниям, чем инбредные линии. Гетерозиготные комбинации аллелей обеспечивают более высокий морфогенетический гомеостаз, чем гомозиготы (Lerner, 1954). В эволюции явно имел место отбор на хорошую комбинационную способность аллелей. (О механизмах поддержания полиморфизма в популяциях см. Kaplin, McGregor, 1972; Северцов, 1990.) В свете этого содержание понятия мутационного груза выглядит слишком большим упрощением. Возможность рекомбинационной репарации, маскирование рецессивных мутаций и взаимодействие аллелей могут намного перекрывать вредные последствия более высокого мутагенеза, связанного с двукратным содержанием ДНК у диплоидов. Таким образом, диплоидность повышает гомеостаз как на уровне организма (более надежный онтогенез), так и на уровне вида: новые мутации могут входить в общий генофонд вида, не нарушая существенно его онтогенетических программ. Преимущество гетерозигот может существенно тормозить эволюцию вида даже при условии высокой приспособленности гомозиготных типов (Michod, 1999). Эволюционные преимущества диплоидии явно не могут быть объяснены с позиции адаптационной эволюции. Есть данные, показывающие, что гаплоидные популяции обладают большей способностью к адаптации в меняющихся условиях среды, чем диплоидные (Zeyl et al., 2003).
Шапероны. Есть еще одно удивительное изобретение эволюции, важность которого не вполне оценена. Это шапероны – класс белков, которые, препятствуя неправильным ассоциациям полипептидных групп, помогают правильному свертыванию или сборке других белков in vivo, но сами не входят в состав зрелых структур (см. обстоятельный обзор Rutherford, 2003). В первую очередь это, конечно, механизм ускорения правильного свертывания полипептидных цепей и сборки белковых структур, замена стохастических процессов (медленных и чреватых ошибками) упорядоченными и целенаправленными. Мало того, шапероны обладают очень важной способностью обеспечивать правильное свертывание и сборку белков при наличии мутаций, которые в отсутствие шаперонов приводили бы к нефункциональным или сильно дефектным структурам. Благодаря этому организмы способны выдерживать большой мутационный груз без утраты жизнеспособного фенотипа.
Типичные шапероны относятся к так называемым белкам теплового шока. Они позволяют организмам успешно преодолевать периоды потенциально летального теплового стресса. Первичная антиэнтропийная и антиэволюционная направленность шаперонов очевидна: шапероны позволяют видам сохранить самоидентичность, несмотря на накопление мутаций. Обратной стороной этого явления оказывается увеличение эволюционного потенциала вида за счет скрытого накопления фенотипически нейтральных мутаций. Полагают, что в периоды внешне-средовых стрессов конкуренция за шапероны со стороны поврежденных белков может приводить к массовой экспрессии ранее скрытых мутаций, что иногда может привести к эволюционному скачку (Ali et al., 1998; Zou et al., 1998). Эти взрывы мутантной экспрессии могут действительно помочь дать ответ на многие трудные вопросы в феноменологии эволюции, но было бы ошибкой рассматривать шапероны как созданный эволюцией механизм ускорения эволюции. Очевидно, что для ныне существующих видов обладание шаперонами есть мощный фактор стабильности и устойчивости к дальнейшей эволюции. Как мы обсуждали выше, для организмов с большим геномом и сложным онтогенезом невозможно достижение такой точности репликации генома, которая сама по себе обеспечивала бы устойчивость вида во времени. Мутации идут неизбежно и неотвратимо, и шапероны помогают видам сохраняться, подавляя фенотипическую экспрессию мутаций, давая время для выработки их более надежной нейтрализации за счет, например, супрессорных дополнительных мутаций. А если какая-то популяция такого стабильного вида, попав в стрессовую ситуацию, не справится с экспрессией ранее скрытых мутаций и породит новый и, возможно, сильно отличный от родительского вид, то это не помешает прежнему виду продолжать свое устойчивое существование во времени.
Фенотипическая пластичность. Наиболее успешно задача сохранения вида неизменным без потери способности адаптироваться к меняющимся условиям жизни решается в эволюции на путях совершенствования регуляторных механизмов. Известно, что чем ниже наследуемость признака, тем ниже эффективность идущего по нему отбора. Уже у бактерий широко распространены ненаследственные адаптации, и чем сложнее организмы, чем выше они стоят на эволюционной лестнице, тем большую роль у них играют ненаследственные модификации, от физиологических адаптаций к внешним условиям и образу жизни до выработки сложных поведенческих реакций на основе жизненного опыта. Пресловутое ненаследование приобретенных признаков, которое так огорчало ламаркистов, – это ведь мощное сопротивление хаосу безрассудной эволюции, вынуждаемой меняющимися обстоятельствами жизни.
Независимость от среды. Дополнительным к рассмотренной выше пластичности фенотипа является другой путь уклонения от эволюции, состоящий в целенаправленной модификации среды обитания, создании искусственных экологических ниш: муравейники, гнезда пчел, плотины бобров, норы грызунов и других животных, гнезда птиц, наконец человеческая культура. Эволюционное значение этих явлений – в оптимизации среды, в приспособлении ее к потребностям вида, а тем самым – в ослаблении давления отбора. Сюда же относится и К-стратегия размножения, когда родители имеют небольшое число детей, но благодаря эффективной заботе о потомстве надежно обеспечивают их выживание. При r-стратегии самки откладывают огромное число яиц, но при этом до взрослого состояния могут доживать единицы на миллион. Очевидно, что K-стратегия в значительной мере выводит потомство из-под давления отбора, обеспечивая выживание всех, кто не погибает от внутреннего несовершенства. К-стратегия, кроме того, позволяет уменьшить количество половых продуктов, а следовательно, и длину зародышевого пути, а следовательно, и конечную частоту мутаций в гаметах. Если на физиологическом уровне фенотипическая пластичность позволяет организмам адаптироваться к меняющимся условиям среды, не меняя генотипа, то на поведенческом уровне организмы могут приспосабливать среду к своим потребностям. Антиэволюционная направленность всех этих усилий природы, от точности репликации ДНК до человеческой цивилизации (с ее комфортом и медициной), очевидна и понятна: выживает, сохраняется лишь то, что не изменяется. Это верно, что вид может оказаться как бы перед выбором: измениться или вымереть. Но это кажущаяся альтернатива. В обоих случаях прежний вид исчезает. Действительным совершенством, к которому влечет эволюция, является «способность выжить, не меняясь».
Роль полового размножения в стабилизации вида обсуждается ниже в разделе II. Консервативная роль полового размножения.
Об адаптивном мутагенезе и эволюции способности к эволюции
В последние годы высказывают сомнения в том, что мутации – просто неизбежное следствие несовершенства механизмов репликации и репарации ДНК, и прокламируют мысль, что возможен отбор на усиление мутагенеза, что высокий уровень мутагенеза может быть адаптивным (Kirschner, Gerhart, 1998; Radman, 1999; Radman et al., 1999, 2000; Giraud et al., 2001; Rattray, Strathern, 2003; Earl, Deem, 2004). При том, что в случайном мутагенезе частота вредных мутаций превышает частоту благоприятных мутаций на 4–5 (Taddei et al., 1997), а то и 6 (Roth et al., 2003) порядков величины, эта мысль не выглядит тривиальной даже при одобрительном отношении ко всякой эволюции. И вопрос не только в том, как могут быть полезны вредные мутации (это еще можно себе представить), а в том, как возможна эволюция способности к эволюции? Это был бы процесс с положительной обратной связью. Динамика таких процессов имеет характер катастрофы. Организмы устроены целесообразно. Их цель – выжить и оставить потомство (то есть сохраниться в будущих поколениях). Эта цель вложена в них эволюцией, и все системы организма служат этой и только этой цели. Служит ли этой цели способность к эволюции? Эволюционировать (то есть меняться) и сохраняться – вещи противоположные. Мысль, что сама эволюция является целью организмов, находится за пределами естественнонаучной парадигмы. Онтогенез осуществляется под управлением генетических программ организма, и его адаптивные реакции на действия среды предусмотрены этими программами; филогенез всегда осуществляется впервые. Это процесс, движимый энтропией, «той силой, что вечно хочет зла». Можно ли предполагать существование механизмов ускорения роста энтропии, чего-то вроде филогенетического апоптоза?
То, что проблема эта всерьез обсуждается на страницах весьма уважаемых изданий (Nature, Genetics, Proc. Natl. Acad. Sci. USA), означает, конечно, что общие гносеологические аргументы не могут противостоять человеческой потребности в чудесах (и сенсациях). Существуют ли, однако, достаточные эмпирические данные или теоретические соображения, позволяющие говорить о механизмах ускорения эволюции, создаваемых в ходе эволюции? У бактерий известны особые высокомутагенные системы синтеза и репарации ДНК, функционирующие только в стрессовых условиях, грозящих организмам гибелью (Friedberg et al., 1995; Drake et al., 1998; Radman, 1999), из которых наиболее изучена система SOS-репарации у кишечной палочки. УФ-облучение или другие воздействия, повреждающие или блокирующие репликацию ДНК, индуцируют экспрессию около 20 генов, продукты которых участвуют в рекомбинации и репарации ДНК. Среди них есть несколько ДНК-полимераз, отличающихся от репликативных полимераз гораздо меньшей точностью репликации. Мирослав Радман, главный адепт эволюции эволюционируемости (evolution of evolvability), предложил называть их ДНК-мутазами (Radman, 1999)1. Индуцируемые системы типа SOS-репарации и ДНК-мутазы найдены и у других микроорганизмов, включая дрожжи.
Интерпретация сторонников адаптивного мутагенеза следующая: организмы, попавшие в стрессовую ситуацию и оказавшиеся перед альтернативой «умереть или измениться», выбирают последнее и включают мощную мутагенную машину в надежде, что хотя бы один из миллионов новых мутантов окажется способным выжить в новых условиях, например в условиях сильной радиации. Подобный альтруизм мне представляется невозможным даже для Homo sapiens, разве что при тоталитарном режиме. Репарационная парадигма выглядит более естественной: SOS-ответ служит репарации повреждений и спасению репликации ДНК ценою пониженной точности, а ДНК-мутазы – это всего лишь нестрогие полимеразы, способные использовать неидеальные матрицы. С помощью такого механизма спастись пытаются все клетки, а не лучшие из лучших. Возможно, что иногда ускоренный мутагенез приводит к появлению организма, приспособленного к среде, вызвавшей стрессовую реакцию, но это не более преднамеренно, чем возникновение сложности при росте энтропии (хотели как хуже, а вышло хорошо). Но и это не очень реалистично. Чтобы, например, стать сверхустойчивым к облучению, организм должен создать сверхмощные системы репарации ДНК, которые как раз и обеспечивают сверхнизкую скорость мутагенеза. Это потруднее, чем вытащить себя из болота за волосы.
Есть еще одна экспериментальная система, в которой бактерии как будто бы обнаруживали «адаптивный мутагенез» (Cairns et al., 1988; Cairns, Foster, 1991; Foster, Cairns, 1992). У lac-мутанта кишечной палочки наблюдался очень высокий уровень реверсий к Lac+ при посеве мутантной культуры на чашки с лактозой в качестве единственного источника углерода. Эти результаты были сначала интерпретированы как способность бактерии направлять мутагенез на селективно важные области генома (Stahl, 1988; Boe, 1990). Позднее была предложена более изощренная модель (Hall, 1992; Rosenberg, 2001), предполагающая существование механизма, который реагирует на стресс индукцией общего (ненаправленного) мутагенеза в небольшой части популяции, подвергнутой стрессу, так сказать созданием передового отряда эволюции. На количественных моделях показана несостоятельность этой гипотезы (Roth et al., 2003), но сторонники направленного мутагенеза не складывают оружие. По-видимому, я поторопился, сказав, что в биологии ламаркизм преодолен. Ламаркизм бессмертен.
Другая группа данных касается систем локального (прицельного) мутагенеза. Лимфоциты, продуцирующие антитела, имеют высокую скорость мутирования вариабельных сегментов иммуноглобулиновых генов, в миллионы раз превышающую нормальный темп мутагенеза благодаря их интенсивной рекомбинации с молчащими псевдогенами (Weil, Reynaud, 1996). Аналогичный механизм используют некоторые паразитические простейшие для прицельного мутагенеза генов, кодирующих поверхностные антигены (Pays, 1989). Гены патогенных бактерий, кодирующие поверхностные антигены, проявляют наследственную повышенную мутабильность (Moxon et al., 1994). Эти гены содержат короткие повторы оснований, на которых нормальные ДНК-полимеразы делают много ошибок (ошибки проскальзывания), что ведет к мутациям типа сдвига фазы. Эти случаи «намеренного мутагенеза» являют собой действительно пример использования мутагенеза в качестве приспособительного признака. Это специализированный вариант приспособительного использования генетического разнообразия, которое в конечном счете не ведет к эволюции вида, а напротив, обеспечивает его стабильность во времени, подобно тому, как полиморфизм популяций позволяет им устойчиво сохраняться в варьирующей среде.
Приводят и другие доводы косвенного характера в пользу отбора на повышенный мутагенез: возможность отбора у бактерий на повышение точности синтеза ДНК (Fijakowska et al., 1993) интерпретируют в пользу отсутствия жесткой эволюции на точность репликации ДНК (Radman et al., 2000); важность генетического разнообразия для выживания интерпретируют в пользу возможности отбора на повышенный мутагенез (Radman, 1999). Но такая интерпретация представляется излишней. Точность репликации у бактерий достаточно высока для устойчивого, неопределенно долгого сохранения вида, и более высокая точность означала бы неоправданные затраты энергии. Что касается генетического разнообразия популяций, то его полезность для долговечности вида достаточно обоснована. Природные популяции, даже бесполые, сильно полиморфны, но надо полагать, что полиморфизм служит экологической пластичности вида, его устойчивости в меняющейся среде. Это не мутационный груз, а богатство генофонда. Множественные аллели уже отобраны на совместимость, на условную полезность и т. п.
В феномене биологической эволюции кроется фундаментальное диалектическое противоречие, очевидное уже в исходной дарвиновской триаде – наследственность, изменчивость, отбор. Изменчивость предполагает несовершенную наследственность. Моя мысль состоит в том, что в ходе эволюции идет преодоление изначального несовершенства наследственности, в пределе – уничтожение изменчивости. Необходимо, однако, различать изменчивость как отражение уровня мутагенеза (неизбежное зло) и разнообразие как богатство генофонда вида, генетическую адаптируемость (как способность к микроэволюции) и приспособленность (адаптированность) как совершенство генофонда, тормозящее дальнейшую эволюцию (Leigh, 1973). На мой взгляд, различение этих понятий позволит избежать логической ошибки, несомненно лежащей в предположении о возможности отбора на повышенную способность к эволюции. Это мое утверждение не означает, что в ходе эволюции объективно не возрастает способность к эволюции. Усложняющаяся биосфера в силу именно своей сложности становится, с одной стороны, всё более уязвимой для энтропии, а с другой стороны, создает всё увеличивающееся разнообразие экологических ниш, что должно вести к увеличению вероятности успешных попыток возникновения и закрепления новых форм. Но в ходе эволюции в геологических масштабах времени сохраняются только те виды, которые достаточно успешно противостоят росту энтропии. Такова диалектика эволюции.
Полезные мутации – вредны. Генетика популяций изучает эволюцию адаптаций, тогда как содержание эволюции – противостояние энтропии. Для победы над энтропией необходимо (помимо всего прочего) устоять против «соблазна приспособленчества», устоять против давления среды, не стать ее отпечатком, остаться самим собой! Только верные наследуют землю! Верные самим себе.
1 Парадоксально, что именно M. Радману принадлежит ведущая роль в открытии пострепликативной репарации у бактерий, доводящей точность репликации ДНК до 10–10 ошибок на пару нуклеотидов (Radman, Wagner, 1986). Когда были открыты неточные ДНК-полимеразы, он принял это известие с восторгом и сказал, что мечтал о них 30 лет (Chicurell, 2001).
Совершенство
В синтетических теориях эволюции в качестве параметра, определяющего направление отбора, используют приспособленность – относительную эффективность размножения особей в популяции. Реальное (не тавтологическое) содержание параметра приспособленности не поддается определению. В любом реальном биотопе, например в пресноводном водоеме, обитают сотни, а то и тысячи видов организмов – бактерии, грибы, водоросли, беспозвоночные животные, рыбы, земноводные, птицы. Все они приспособлены к данному биотопу. И если все эти виды продолжают эволюционировать, то общее направление эволюции, очевидно, будет определяться собственной историей вида, а не средой обитания (Brooks, Wiley, 1986). Разумеется, при этом виду придется платить нечто за сохранение приспособленности к среде, приспосабливаться, но по-своему. При таком понимании направленности эволюции можно и согласиться со сторонниками ортогенеза. Может ли эволюция создавать всё что угодно, как это молчаливо предполагается синтетической теорией эволюции?

«Адам и Ева в Земном раю». Картина Венчеслао Петера (Wenceslao Peter; 1742, Богемия – 1829, Рим), Пинакотека, Ватикан. Согласно концепциям Естественной истории (XVI-XVIII век), Природа – это идеально сбалансированная устойчивая система, задуманная и поддерживаемая Творцом. В этом мире, конечно, нет никакой эволюции! На полотне узнаваемы десятки видов птиц и млекопитающих, мирно сосуществующих. Венчеслао Петер был известен прежде всего как анималист. (Изображение с открытки, изданной Ватиканскими музеями)
Сам Дарвин вкладывал в понятие приспособленности представление о совершенной конструкции. Хорошо ощущаемое интуитивно, понятие совершенства ускользает от определения. Описанный здесь взгляд на эволюцию позволяет, мне кажется, внести некоторую ясность в проблему совершенства и прогресса в биологии и связать ее с обсуждавшимся выше вопросом, что же отбирает отбор. Что совершеннее – семейство сложноцветных, отряд перепончатокрылых или отряд приматов? Пчела или человек? Или, может быть, цианобактерия, сотворяющая всю органику из света, воды и минеральных солей? Критерий близости к человеку не выглядит удовлетворительным, во всяком случае как единственный. Одним из критериев совершенства может быть долговечность, способность к выживанию без изменений. Это не абсолютное совершенство, а только совершенство в своем роде, но это важный и содержательный критерий – способность противостоять энтропии.
Что касается приспособленности, то никто не рассматривает ее как способность выживать не меняясь. Напротив, приспособленность, по определению, коррелирует со скоростью эволюции. Интерес популяционной генетики сосредоточен на изменениях, ибо ее предмет – эволюция адаптаций. Но изменения сами по себе есть энтропия, а шагом к совершенству является только такое изменение, которое препятствует дальнейшим изменениям. Эволюционная история обнаруживает градиент структурной сложности. Поскольку сложность сама по себе означает большую способность к деструкции, эволюционное значение имеет только организованная сложность. Стабильность сложной системы критически зависит от установления специфических, неслучайных связей между ее частями (Saunders, Ho, 1976), которые обеспечивают ее функционирование. Эти связи являются результатом естественного отбора и не могут быть выведены из свойств компонентов системы (Wicken, 1979). Единицей естественного отбора должна быть, следовательно, организация как целое. Каждый шаг в направлении приспособленности к условиям среды и каждый шаг к увеличению структурной сложности должны быть компенсированы общим изменением организации как системы противостояния росту энтропии. Не пытаясь дать определение абсолютному совершенству, можно указать направление, в котором действует эволюция, «стрелу совершенства»: возрастание организованной сложности.
В период противостояния генетики и дарвинизма в начале ХХ века для генетиков было характерно представление об идеальной особи, совершенном типе, а изменчивость рассматривали как результат ошибок (мутаций), связанных с несовершенством механизмов наследственности – типологическое мышление, ведущее свое начало от Платона. Генетики увлеклись евгеникой, но национальная трагедия Германии, попытавшейся на практике осуществить освобождение человечества от несовершенных его членов, помогла оценить достоинства популяционного мышления. Рассмотренные выше преимущества полиморфных популяций в сравнении с чистыми линиями и известные негативные последствия пресловутой зеленой революции привели к пониманию, что совершенство вида в его разнообразии, богатстве его генофонда. Это разнообразие вносит существенный вклад в гомеостаз вида, в его способность противостоять энтропии.
Генетический «замысел» зиготы
Построение многоклеточного тела можно рассматривать как деятельность зиготы, направленную на создание оптимальных условий для сохранения и размножения клеток зародышевого пути. Курица – это способ, которым яйцо порождает яйцо. При таком взгляде вся эволюция сводится к эволюции зародышевого пути, непрерывной зародышевой плазмы, побочным результатом которой оказывается эволюция многоклеточных организмов, более высокоорганизованных монад2, чем клетки. Организмы различаются по уровню организации, тогда как генеративные клетки всех организмов являются одноклеточными и не могут быть представлены иерархически. Если это так, если, например, зигота человека мало отличается от зиготы дождевого червя, то как возникает конечная разница между человеком и червем? Это ведь должно быть предопределено в зиготах. Где источник новой информации, появляющейся в онтогенезе? Редукционисты склонны думать, что в онтогенезе никакой новой информации не возникает, что идет просто дублирование и переформулирование информации, содержащейся в хромосомах зиготы. П. Б. Медавар справедливо охарактеризовал такую точку зрения как самое беспомощное описание развития. О громадной разнице в информационном содержании между зиготой и взрослым организмом см., например, в работе (Riedl, 1978).
Онтогенез многоклеточного организма может быть представлен как разветвляющаяся последовательность необратимых трансформаций, каждый этап которой сопровождается появлением новой информации, не содержавшейся в структурах предшествующего этапа в явном виде и не выводимой из них однозначно в соответствии с каким бы то ни было алгоритмом. Новое содержится в предшествующем в качестве возможного. Эту скрытую (потенциальную) информацию зиготы я обозначил здесь термином «генетический замысел». Несмотря на креационистское звучание, он правильно отражает объективное содержание данной информационной категории, не имеющей аналогии в неживых системах, но весьма сходной с тем, что мы вообще вкладываем в понятие «замысел». Это не план, не проект, не модель. Это то, что нуждается в творческом воплощении.
Генетическим замыслом зиготы дождевого червя является дождевой червь, а генетическим замыслом зиготы человека является человек. Содержание генетического замысла последовательно, поэтапно развертывается в онтогенезе. Наглядные примеры реализации генетического замысла дают процессы самосборки субклеточных структур. В клетке существует группа генов, кодирующих рибосомные белки. После транскрипции этих генов и трансляции оказывается, что линейные последовательности аминокислот (полипептиды) содержат информацию для свертывания их в трехмерную белковую структуру. Далее оказывается, что эти трехмерные молекулы обладают специфическим сродством друг к другу и к молекулам рибосомных РНК. Они определенным образом соединяются между собой, образуя рибосому, молекулярную машину, предназначенную для синтеза полипептидов на матрице информационной РНК. Процесс сборки рибосомы идет самопроизвольно и необратимо, с созданием сложности, наделенной функцией (организации). Самосборка привела к появлению новой информации, не содержавшейся ни в ДНК, ни в белковых молекулах. Точно так же идет самосборка других сложных полиферментных комплексов (молекулярных машин), осуществляющих те или иные молекулярно-биологические процессы. Мы говорим, что белки обладают специфическим сродством друг к другу и взаимодействуют друг с другом и с другими компонентами клетки. Мы говорим также, что белки «узнают» друг друга. Это знание – результат эволюции организма как целого, включающей отбор на способность генных продуктов образовывать функциональные сложные структуры. Эта пригнанность, это сродство белков друг к другу не содержатся в явном виде в последовательности нуклеотидов в генах, кодирующих эти белки. О существовании этого сродства мы можем судить только a posteriori.
Можно возразить, что мы можем по последовательности нуклеотидов в гене судить о трехмерной структуре кодируемого белка, а затем и о возможном сродстве между белками. Однако фундаментальная теория, способная по первичной последовательности аминокислот в полипептиде предсказывать трехмерную структуру белка, невозможна. Теории свертывания полипептидов являются эмпирическими. Переход же отсюда к взаимодействию белков будет связан с еще большей неопределенностью. Следующие слои онтогенеза, идущие через взаимодействие клеток, зародышевых листков, органов, находятся уже так далеко от ДНК зиготы, что их предсказуемость по ДНК находится за рамками научной фантастики. Таким образом, информация, определяющая целостное развитие, хотя и содержится в зиготе, содержится в ней как потенция и предварительному декодированию недоступна.
Наиболее сильный аргумент против сводимости биологии к физико-химии представляет сама последовательность ДНК, которая не может быть выведена из физико-химии нуклеотидов. Напротив, эта невыводимость — необходимое условие, позволяющее ДНК содержать генетическую информацию, содержать код. Последовательность – это зафиксированная отбором случайность. Нельзя предсказать текст литературного произведения по алфавиту. Между прочим, не следует заблуждаться относительно нашей способности читать генетический код. Мы это умеем делать только потому, что код оказался универсальным. Мы его узнали эмпирически. Связать структуру триплетов ДНК с определенными аминокислотами, исходя из физико-химических свойств триплетов и аминокислот, невозможно, ибо этой связи нет. Связь эта символическая. Мы читаем генетические тексты только потому, что все организмы Земли «говорят» на одном языке. Когда клетка сама читает генетические тексты, переводя их сначала в тексты РНК, а затем и в аминокислотные последовательности, она использует аппараты, основанные на физико-химических взаимодействиях, но роль их только служебная. Все сущностные процессы биологии имеют характер символичности. Ни физико-химия вообще, ни энергетика в частности, не определяют живое. Хотя живое во всём этом нуждается, определяющими являются смысловые составляющие, не физико-химическая причинность, а смысловые сигналы: «Вдруг слабым манием руки на русских двинул он полки». И никакого «causa equat effectum». Общая теория биологии должна быть семантической, а не физико-химической. Организм гораздо ближе к компьютеру, чем к химическому ферментеру.
Самосборка и самоорганизация являются тем механизмом, который реализует, материализует, воплощает генетический замысел. Ферменты, другие макромолекулы априорно знают друг о друге и благодаря этому способны к организации в функциональные структуры. Совершенствование способности к самоорганизации и есть главное содержание биологической эволюции. В генетическом замысле потенциально содержится не только знание частей организма друг о друге, но и знание о внешней среде. Все организмы умеют распознавать в окружающем мире нужные им компоненты, так же как и компоненты вредные или опасные. Ферменты знают свои субстраты и умеют с ними обращаться, животные распознают в окружающей среде пищу и врагов. Для всего этого в ДНК зиготы не содержится прямых инструкций. Они создаются в ходе онтогенеза. Бесполезно искать в ДНК зиготы закодированные инстинкты или музыкальный талант – они возникают на последних этапах самосборки организма. Концепция генетического замысла позволяет понять, каким образом в ходе индивидуального развития идет наращивание сложности и организованности в кажущемся противоречии со вторым законом термодинамики. Теории термодинамики разрабатывали для характеристики поведения атомов, то есть монад, лишенных разума и знаний. При этом поведение монад определяется правилами теории вероятности. Когда же мы переходим к поведению биологических монад (генов, белков, клеток, организмов и т. д.), мы не имеем права игнорировать наличие у них способности к целенаправленному поведению. Понятию целенаправленности я придаю здесь достаточно широкий смысл, не обязательно включающий осознанное поведение, а только поведение, ведущее к определенному, заданному результату. Например, поведение рибосомных белков и рибосомных РНК таково, что оно всегда приводит к образованию рибосомы (в адекватной среде).
Процесс развертывания генетического замысла – творческий процесс, поскольку включает в себя оба необходимых компонента всякого творчества: случайный перебор вариантов и отбор правильного варианта. По своему содержанию генетический замысел не идентичен ценности информации. Ценность информации предполагает возможность ее однозначного декодирования знающим приемником, аналогично переводу текста на другой язык. При этом новая информация не создается. В онтогенезе же идет сотворение, воплощение в реальность того, что существовало лишь как возможное. Замысел – это третий, вернее первый, компонент всякого творчества. Замысел не расшифровывается, не декодируется, а эмпирически (методом проб и ошибок) воплощается.
Когда появились данные по последовательности нуклеотидов в ДНК человека и других организмов, мы были удивлены, как мало человек отличается не только от шимпанзе, но и от мухи и даже от червя, если судить по ДНК. Но замысел не связан напрямую ни с объемом генетических текстов, ни с количеством глав и параграфов в этих текстах. Идея невыводимости высшего уровня организации из предшествующего уровня более низкого ранга не нова. Например, Е. Г. Мерсер (Merser, 1981) утверждает, что действующие принципы организации более высокого ранга не обязательно являются производными свойств компонентов, которые они контролируют, или их внутренних действующих принципов. Адаптационные механизмы и организационные ограничения живых организмов, возникая случайно, не могут быть выведены из какого-либо общего принципа или закона и могут быть поняты только в контексте их истории. Сходные воззрения о несводимости граничных условий биологических систем к физике и химии высказывал М. Полани (Polanyi, 1968). Очевидно, что ДНК не является алгоритмом организма, его сжатой формулой. Организм есть организация, его информация обладает, следовательно, свойством несжимаемости (Denbigh, 1975).
В биологической эволюции определяющими становятся изменения генетического замысла зиготы. Эволюция зародышевой плазмы есть эволюция граничных условий онтогенеза. Граничные условия – это тоже законы, но не фундаментальные законы физики, а законы (правила), наложенные эволюцией, которые больше похожи на законы юридические, чем на законы физики. Если законы физики можно считать внутренними свойствами вещества и энергии, то граничные условия – это внешние законы. В машинах и организмах вещество и энергия, продолжая подчиняться законам физики, оказываются также в подчинении внешним законам. Внешние законы действуют и в неодушевленном мире: река течет, подчиняясь внешним ограничениям русла, но только машины и организмы имеют «разумные» граничные условия, придающие конструкции целесообразность. Эволюция организмов идет через отбор граничных условий на эффективность использования энергии и вещества для созидания. Такая направленность эволюции автоматически рождается из того факта, что выживает только то, что выживает. Произошло землетрясение, и сохранились только те здания, которые устояли. Это, конечно, тавтология, но только до тех пор, пока мы не интересуемся вопросом, почему одни здания разрушились, а другие устояли. Механизмы жизнеспособности могут быть самыми разными. Организмы размножаются, и удачные конструкции воспроизводятся.
Законы, управляющие онтогенезом, это не физико-химические запреты, а семантические правила. Реализуются эти правила физико-химическими взаимодействиями, ведущими к самосборке. На каждом этапе самосборки возникают новые конфигурации, обеспечивающие возможность новых взаимодействий. Онтогенез можно сравнить с многомерной детской головоломкой, в которой каждый этап сборки приводит к созданию новых конфигураций, отсутствующих среди первоначальных деталей. То есть генетический замысел – это свернутая информация о многомерной организации, где роль измерений играют уровни организации: генетический, биохимический, морфологический, физиологический, нервно-психический, интеллектуальный, поведенческий, духовный.
Продолжая старинный спор о преформизме и эпигенезе, я говорю в пользу эпигенеза: в ходе онтогенеза информация творится заново, эпигенетически. Но на место жизненной силы я ставлю менее таинственное представление о генетическом замысле. Впрочем, если угодно, эту свернутую информацию можно трактовать как информационный эквивалент гомункулюса преформистов.
Онтогенез есть эмпирическое воплощение генетического замысла об организме. Эпитет «эмпирическое» призван подчеркнуть принципиальную непредсказуемость каждого этапа онтогенеза на основании свойств компонентов предыдущего этапа. Эта непредсказуемость несет в себе большую мировоззренческую нагрузку. Из нее следует уникальность и неповторимость каждого индивидуума и космическая значимость каждой жизни.
Филогенез многоклеточного вида формально похож на онтогенез индивида. Он тоже представляет собой последовательную смену этапов от одноклеточности до современной высокоорганизованной формы. Насколько сущностна эта аналогия? Можно ли говорить о филогенетическом замысле? Положительный ответ на этот вопрос увел бы нас за пределы естественнонаучной парадигмы. Мы ограничимся философской констатацией, что современное состояние Вселенной наличествовало в новорожденной Вселенной в качестве возможного. Интересно, что само слово «эволюция», придуманное Шарлем Бонне, означает развертывание (e-volvo — развертывать). Бонне просто перенес представление об эмбриогенезе на филогенез. Взгляд на мир как на гигантский организм, наделенный импульсом к прогрессивному развитию, высказывали и Платон, и Августин, и Кант, и Гёте. Принципиальное различие между филогенезом и онтогенезом состоит, однако, в том, что филогенез осуществляется впервые, тогда как развитие индивидуальных организмов – это варианты воспроизведения истории вида, предопределенные генетическим замыслом. Действительно новое возникает в филогенезе. Оно возникает как изменение в зародышевой плазме. Пресловутая антиномия «яйцо или курица?» – недоразумение. Конечно, яйцо. Если изменение в замысле оказалось удачным, то есть было санкционировано естественным отбором, оно унаследуется в виде измененного генетического замысла. Таков механизм биогенетического закона – онтогенез повторяет филогенез.
Образно говоря, генетический замысел представляет собой подобие туго скрученной пружины, которая развертывается в ходе онтогенеза. Это развертывание, это наращивание организованной сложности производит впечатление чуда.
Разгадка – в происхождении генетического замысла. Далеко не каждое случайное изменение в генотипе передается потомкам, а только такое, которое реализуется в жизнеспособный организм. То, что мы видим в окружающей нас живой природе, – это ничтожная выборка удачных вариантов из мириад других, менее удачных. За нас, ныне живущих, заплачено очень дорого. Мы – редкая случайность, зафиксированная отбором. Такой отбор, отбор редких организаций со свойством выживания – организаций, противостоящих энтропии, есть Демон Максвелла или Бог Дарвина. Глобально, с учетом всей истории биосферы, второй закон не нарушен, но локально идет сотворение маловероятного мира.
Как обстоит дело с энтропией эмбриогенеза? Эмбриогенез – необратимый процесс, а необратимость предполагает рост энтропии. Брукс и Вили (Brooks, Wiley, 1986) считают эмбриогенез таким же энтропийным процессом, как и филогенез. В самом деле, структурная энтропия взрослого организма, состоящего из сотен миллиардов клеток, намного больше структурной энтропии зиготы. Для объяснения очевидного возрастания информации в онтогенезе они привлекают идею Дэвида Лэйзера, что источником новой информации является рост разности между максимально возможной энтропией и действительной энтропией организма, которая растет с меньшей скоростью (Layzer, 1975, 1977). Эту разность можно оценить, сравнив действительную энтропию беспорядочного нагромождения сотен миллиардов клеток с действительной энтропией живого организма. Рост энтропии – необходимое условие всякого движения, однако не рост энтропии, а генетический замысел каузально определяет направление эмбриогенеза. Здесь исторические ограничения выражены в гораздо большей степени, чем в филогенезе. Эмбриогенез довольно жестко детерминирован в пределах нормы реакции генотипа.
Если наблюдать за строительством здания со стороны и ничего при этом не знать ни об архитекторе, ни о прорабах, ни о каменщиках, то постройка здания покажется процессом совершенно невероятным. Но подумав, мы можем догадаться и о замысле архитектора, и о мастерстве рабочих, и об их руководителях, а может быть, и о совершенно особой необратимости – необратимости управляемых процессов.
2Не найдя подходящего современного русского слова, я отваживаюсь использовать этот термин Лейбница в качестве универсального названия для индивидуализированных элементов и систем разного уровня организации.